
A complex life cycle involving more than one host is a common feature of parasitic animals. This bestows many advantages but poses a problem – how to get from one host to the other. Some parasites have life cycle stages that are motile and actively seek out hosts, others use hosts (vectors) that deliver them and many use hosts that are linked by a food chain and are transmitted to the next host when their current one is eaten. This is known as trophic transmission.
Many trophically transmitted parasites have tricks up their sleeves to enhance the chance that their hosts are eaten; but only once they have developed to a stage capable of infecting the next host. For example, when infected, their host may lose its predator avoidance behaviour and even behave in a way that attracts predators.
Spiny-headed worms
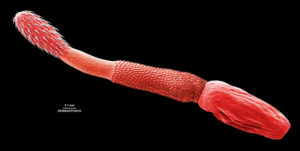
One group of parasites, the spiny-headed worms or acanthocephalans, are known to cause behavioural changes such as these to occur in their invertebrate hosts. As adults, acanthocephalans inhabit the guts of a variety of vertebrates, punching their spiny headed proboscis through the gut wall to anchor themselves.
Eggs pass out into the environment and, if eaten by a species that is prey for the vertebrate host, there is a chance that the acanthocephalan will complete its life cycle. One such example is Polymorphous minutus.
Polymorphous minutus
If a P. minutus egg is ingested by a shrimp it will hatch in its gut and burrow through into the body cavity and develop via an acanthella stage into a barrel shaped stage, the cystacanth.

If the infected shrimp is eaten by a waterfowl the cystacanth will develop into a mature worm in the bird intestine.
The effects of infection
Shrimps normally dive to the bottom of a pond or stream if they sense the presence of a predator. Those that contain an acanthella also have a strong positive geotaxis, a behaviour that would protect the developing parasite. However, once the parasite becomes a cystacanth there is a dramatic change in shrimp behaviour. They display negative geotaxis, swimming to the surface and clinging to floating vegetation, thus becoming exposed to feeding waterfowl (see previous BugBitten blog).
A study of this infection in the shrimp Gammarus roeseli suggests that low oxygen availability and excretory products associated with parasite anaerobic respiration may be responsible for these changes.
In addition to exhibiting behavioural changes, none of the eggs of female shrimps infected with this parasite develop and male reproductive success is reduced.
Underlying mechanisms
Both parasite-induced reproductive disturbances and altered behavioural traits are commonly seen in infected invertebrates but have mostly been investigated separately. Mechanisms underlying these effects are still poorly understood. In particular, we do not know if different phenotypic alterations are initiated by a single disruption of host physiology or if multiple causes are involved.
Colleagues at the Université Bourgogne Franche-Comté, France, have investigated this question by studying P. minutus infections in the shrimp Gammarus pulex.
Looking for links
The study looked at behavioural changes and sterility with respect to relative time courses, and any correlations between them. Shrimps were collected from the wild. Using a microscope, parasites were observed through the translucent cuticle of the shrimp, their developmental stage noted, and the sex of the shrimp determined. As the collecting season progressed from February to April the number of infections with acanthellae declined from 63% to 0% as they matured into cystacanths.
Behavioural changes
Individual infected and non-infected shrimps were monitored swimming in long glass columns to determine their geotaxis. Shrimps with similar developmental stages were compared with uninfected shrimps. In all cases males and females behaved in the same way and there was no difference in the geotaxis score over time for uninfected shrimps.
The story for infected shrimps was very different and confirmed the observations of several previous studies. Shrimps infected with the early, acanthella stage had a stronger positive geotaxis than uninfected ones. Those with a cystacanth had negative geotaxis; behaviour that would bring them in the proximity of feeding wildfowl such as mallards. This shift in behaviour occurred over several days.
Reproductive potential

Male shrimps guard egg- bearing females by clasping them from behind with special claws and carry them for several days pre-copulation. Mating behaviour was assessed by placing an infected male with an uninfected female that was receptive to mating and allowing it one hour in which to grasp her. Infected males showed a very significant decline in this mating behaviour.
Male reproductive traits were assessed by examining the size of these claws and by calculating sperm reserves. Although claw size was not affected by the presence of a cystacanth, sperm reserves were significantly increased at this infection stage, but not earlier. Sperm were probably accumulating if female insemination was not occurring.
Female fecundity was assessed by counting the number of eggs or embryos present in the brood pouch. Infection did not cause a difference in the number of eggs, however, infected females, regardless of the stage of infection, had virtually zero developing eggs. The authors use the term castration to describe this. Although strictly incorrect, as there is no irreversible destruction of reproductive tissue involved here, the term parasitic castration is in general use to describe infections that have a negative impact on host reproduction (see my discussion in Advances in Parasitolgy, 29)
Correlations
Both the lack of differences in temporal changes in behaviour and castration and a correlation between intensity of behavioural changes and intensity of castration did not point to a single mechanistic cause for these phenomena. However, the authors point out that products excreted by the parasite or from a host immune response may accumulate over time, and traits may respond differently to different concentrations of these products. Taxis behaviours may even respond in a bi-phasic way to different concentrations. They have not yet ruled out a common mechanism to explain a link between different phenotypic alterations.
Further studies of this and other models are clearly required, as identifying these mechanisms has important bearings on the whole debate concerning host manipulation by parasites.

Comments